![]()
Structural alteration in chromosome
STRUCTURAL ALTERATIONS IN CHROMOSOMES
- The expression and inheritance of the sum total of characters of an organism is determined by the number and sequence of genes of its cells.
- The changes in architecture and morphology of chromosomes do not involve changes in the number of chromosomes, but result from changes in the number or sequence of genes of chromosomes.
- These changes are collectively known as chromosomal aberrations and were first analyzed by Muller (1928) in Drosophila and by Barbara McClintock (1930) in Zea-mays.
- Cytological observations along with genetic studies have resulted in the identification of following types of changes in the number or arrangement of genes:
- All types of chromosomal aberrations are caused by a break in the chromosome or its chromatid.
- When broken ends rejoin to restore the original chromosome structure and gene sequence, there is no change and restitution occurs.
- Aberrations occur when broken ends are lost or get joined to a wrong chromosome or in a wrong order.
- The number of breaks, their location, and the pattern in which broken ends get joined determines the type of chromosomal aberration.
- Their study is important because they occur in nature quite frequently and have been instrumental in the evolution of many species of plants and animals.
- Their frequency can be increased with the help of ionizing radiations or certain chemical agents.
A. Changes Involving Number of Genes
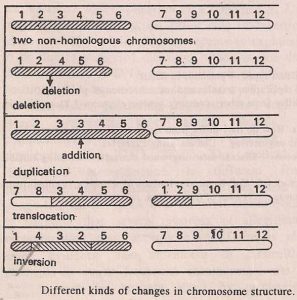
i) Deletion or deficiency
It signifies the loss or absence of a section of a chromosome and may involve one or more genes.
ii) Duplication.
It represents a gain in chromosomal material and in this case a part of a chromosome is present in excess of the normal amount. Thus one or more genes are present in more than two doses.
B Changes Involving Arrangement of Genes
i) Inversion.
In this process a section of a chromosome gets inverted or rotates by 180° on its own axis. This changes the relative positions of genes on a chromosome, but not their number.
ii) Translocation.
It involves the transfer of a part of one chromosome to a nonhomologous one, thereby changing the relative positions of genes but not their number.
Deletion or Deficiency
- If a section of chromosome gets broken and detached, some genes are lost from the chromosome.
- Acentric fragments are either digested by nucleases or fail to move to either of the poles during cell division.
- Usually they get excluded from the daughter cells.
- Sometimes they move towards one end of the cell, just by chance, and get included in one of the two daughter cells.
- In either case a population of cells arises which is deficient for a few genes.
- If gametes arise from deficient cells, deletion is transmitted to the next generation.
- These are the following type
Terminal Deletion
- In this type of Deletion a terminal section of chromosome is absent,
- Only one break is necessary for terminal deletion,
- Broken acentric fragments of chromosomes, which are in the process of being digested, sometimes appear as small chromatin bodies in cells and are known as micronuclei.
Intercalary Deletion
- In this type of deletion an intermediate section or portion of chromosome is lost.
- for intercalary deletion two breaks (one on either end of the deleted region) are essential.
- A break for intercalary deletion results into three pieces of chromosome, the middle one of which is lost and the other two get joined again.

- Experimental proof for deletion was obtained by Bridges (1916-1919) who studied the inheritance pattern of sex linked lethal characters which had arisen spontaneously in a population of fruit-flies.
- Deletion can be recognized by distortions of chromosomes during meiosis pairing of homologous chromosomes or during somatic pairing in specialized tissues like salivary glands of Drosophila.
- Due to a terminal deletion one of the paired chromosomes appears to be much longer than the other.
- This can be seen during pachytene in maize and in salivary gland chromosomes of Drosophila.
- Due to an intercalary deletion the normal chromosome forms a loop near the deficient region of its homologous, because only identical regions pair with each other.
- Inheritance patterns of genes of deleted regions and cytological studies of pairing between normal and deleted chromosomes have helped a lot in finding out the relative positions of genes on chromosomes.
- Thus, deletions have helped in constructing and verifying linkage maps of a variety of organisms like maize, Drosophila, bacteriophages etc.
Duplication
- The presence of a part of a chromosome in excess of the normal complement is known as duplication.
- A broken section of a chromosome attaches itself to a normal homologous or non homologous chromosome or in the presence of a centromere behaves like an independent chromosome and gets included in an otherwise normal nucleus.
- Consequently some genes are present in a cell in more than two doses.
- Depending on the mode of joining of the duplicated region to a chromosome or its independent existence,
- duplications can be of the following types:
1) Extra-chromosomal duplication.
In the presence of a centromere the duplicated part of a chromosome may behave as an independent chromosome.
2) Tandem duplicated.
In this case the duplicated region situated just by the side of the normal corresponding section of the chromosome and the sequences of genes are the same in the normal and duplicated regions.
For example, if the sequence of genes in a chromosome is ABC.DEFGH (full stop represents the centromere) and if the section containing the genes DEF is duplicated, the sequence of genes in tandem duplication will be ABC.DEFDEFGH.

3) Reverse tandem duplication.
In this case the sequence of genes in the duplicated section of a chromosome is just the reverse of a normal sequence. In the above example, therefore, the sequence of genes as a result of reverse tandem duplication will be ABC.DEFFEDGH.
4) Displaced duplication.
In this case the duplicated section is not adjacent to the normal section Depending on whether the duplicated portion is on the same side of the centromere as the original section or on the other side; displaced duplication can be homobrachial or heterobrachial:
homobrachial duplication = ABC.DEFGDEFH.
heterobrachial duplication = ADEFBC.DEFGH.
5) Transposed duplication.
- In this case the duplicated section is attached to a non homologous chromosome.
- If ABC.DEFGH and LMNOPQ.RST represent the genes sequences of two non homologous chromosomes, a transposed duplication will result into chromosomes with gene sequences ABC.DEFGH and LMNDEFOPQ.RST.
- The duplicated region can be transposed to a non homologous chromosome interstitially or terminally.
- Duplications involving various regions of the X chromosome in Drosophila have been studied in detail.
- Flies heterozygous for the Bar (B) gene have rod shaped or rectangular eyes instead of normal round eyes.
- Flies homozygous for this gene have very small eyes.
- In 1936 Muller and Bridges found independently of each other that bar eyes are caused due to a duplication of the region of the X chromosome.
- A reduplication of this region results into Ultrabar or doublebar or very small eyes.


- Duplications and reduplications of the Bar region can be very easily seen in the salivary gland chromosomes.
- As the number of Bar genes increases, the eyes become smaller.
- Duplications, reduplications and deletions may arise due to unequal cross-overs.
- Like deletions duplications also result into unequal or looped out configurations at the time of pairing of homologous chromosomes. Crossing –over in reverse tandem duplication results into a dicentric chromosome.
- This can frequently seen in chromosomes of maize.
- Irrespective of whether sister or non-sister chromatids are involved in a cross-over, dicentric chromosomes are produced.
- As the two centromeres move towards two different poles, a chromosome “bridge” is formed which later on breaks at any point along the bridge.
- The broken ends become “sticky” and a replication of the chromosome produces two sister chromatids which are joined together due to their sticky ends.
- Therefore, the bridge-breakage-fusion cycle goes on indefinitely.
- In sporophytic tissues of plants however, sticky ends are not found.
- Duplication has played a great role in evolution.
- It is a means of increasing the number of genes in a cell so that different copies of the same gene may change in different directions without disturbing the normal functions of an organism.
- It has been suggested that all cases in which different gene pairs affect the same character (e.g. multiple factors, complementary genes, etc.) arose initially as duplications of single genes .
- More evolved organisms have been found to have more repetitions of DNA sequences.
Inversion
- Inversion involves a rotation of a part of a chromosome or a set of genes by 180° on its own axis.
- Breakage and reunion is essential for inversion to occur and the net result is neither a gain or loss in the genetic material but simple a rearrangement of the gene sequence.
- Inversion that include the centromere are known as pericentric (around the centromere), whereas those which do not involve the centromere are known as paracentric ones.

- If the normal sequence of gene in a chromosome is ABC.DEFG, the sequence in paracentric and pericentric inversions will be ABC.DGFE and AED.CBFG, respectively.
- In organisms which are homozygous for inversions, zygotene and pachytene is normal because of similarity of abnormalities in the homologous chromosomes.
- But heterozygous inversions result into looped configurations during pachytene.
- If the heterozygous inversion is small enough, the opposing inverted regions fail to pair and crossing-over is suppressed in this part of the chromosome.
- A crossing-over in the inverted region of a heterozygous paracentric inversion results into a chromosome with two centromeres and an acentric fragment.
- As the two centromere of the dicentric chromosome move towards two opposite poles, a chromatid bridge is formed at anaphase I.
- The acentric fragment is not attached to the spindle, lies free in the cytoplasm and is not disjuncted properly.
- Thus meiotic separations are usually abnormal.
- In case meiotic separation is normal, four types of gametes are formed – one with a normal gene sequence, second with an inverted gene sequence, third with a dicentric chromosome and duplication of some genes, and fourth with an acentric chromosome and deletion of some genes.
- The latter two types of gametes are usually not viable, with the result that heterozygotes for paracentric inversions are highly sterile and only parent-like progeny are produced.
- In other words, crossing-over is apparently suppressed due to paracentric inversion.
- Dicentric bridge formed in maize female tissue as a result of such crossing-over results into a bridge-breakage-fusion cycle.
- Pollen grains show a high degree of sterility.
- Multiple cross-overs reduced the suppression effect, especially with respect to those genes between which two (or even number of) cross-overs occur, because their result is the same as no cross-over (only if the same two strands are involved in even number of exchanges).

- Crossing-over in a heterozygous pericentric inversion does not result into a chromatid bridge, but results into deletions and duplications in the gametes.
- Therefore, pericentric inversions also apparently suppress crossing-over.
- Although heterozygotes for pericentric inversions produce a significant proportion of inviable gametes and unbalanced zygotes as a rule,
- Drosophila is an exception because in the females there is reduced paring and consequent reduced crossing-over in the inverted region between the homologous chromosomes.
- Pericentric inversions involving unequal arms result into drastic changes in the morphology of chromosomes.
- For example, metacentric (V-shaped) chromosomes can be transformed into acentric (rod-shaped) ones or vice-versa.
- Fertility of inversion of homozygote and sterility of inversion heterozygote leads to the establishment of two groups which are mutually fertile but do not breed well with the rest of species.
- Thus, two varieties are established which evolve in different directions and later become reproductively isolated species.
- There is plenty of cytological evidence to prove that such evolutionary mechanisms have and are operating in Drosophila and a number of other organisms.
- Inversion has been useful in establishing and maintaining a heterozygous condition, because in inversion heterozygotes crossing-over is suppressed and only parental progeny are produced.
- Recessive lethal can be added advantage because heterozygotes for them will be viable but homozygote nonviable.
- Quite a few strains of Drosophila are routinely maintained with the help of lethal genes and inversion heterozygosity.
Translocation
- Translocation involves the transfer of a part of a chromosome or a set of genes to a non-homologous chromosome.
- It results into a change in the sequence and position of the genes but not their quantity, means changes involving arrangement of genes.
- An important type of translocation having evolutionary significance is known as reciprocal translocation or segmental interchanges which involve mutual exchange of chromosome segments between two pairs of non-homologous chromosomes…

Related posts:
 CHROMATIN EUCHROMATIN AND HETEROCHROMATIN
CHROMATIN EUCHROMATIN AND HETEROCHROMATIN
 ANEUPLOIDY
ANEUPLOIDY
 MOLECULAR ORGANISATION OF CENTROMERE AND TELOMERE
MOLECULAR ORGANISATION OF CENTROMERE AND TELOMERE
 NUCLEOLUS AND RIBOSOMAL RNA GENES
NUCLEOLUS AND RIBOSOMAL RNA GENES
 Molecular Mechanism of Chromosomal Pairing
Molecular Mechanism of Chromosomal Pairing
 Structure of Chromosome
Structure of Chromosome
 Specialized Type of Chromosome
Specialized Type of Chromosome
 NUCLEOSOME ORGANIZATION
NUCLEOSOME ORGANIZATION
 PACKAGING OF DNA
PACKAGING OF DNA


