Initiation of Cancer at cellular level
Cancer
- A cancer is an uncontrolled proliferation of cells. In some the rate is fast; in others, slow; but in all cancers the cells never stop dividing.
- This distinguishes cancers – malign tumors or malignancies – from benign growths like moles where their cells eventually stop dividing.
- Cancers are clones.
- No matter how many trillions of cells are present in the cancer, they are all descended from a single ancestral cell. Evidence:
- Although normal tissues of a woman are a mosaic of cells in which one X chromosome or the other has been inactivated, all her tumor cells – even if from multiple sites – have the same X chromosome inactivated.
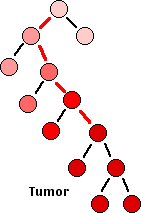
- Cancers begin as a primary tumor.
- At some point, however, cells break away from the primary tumor and – traveling in blood and lymph – establish metastases in other locations of the body.
- Metastasis is what usually kills the patient.
- Cancer cells are usually less differentiated than the normal cells of the tissue where they arose.
- Many people feel that this reflects a process of dedifferentiation, but I doubt it.
- Rather, evidence is accumulating that cancers arise in precursor cells – stem cells or “progenitor cells” – of the tissue: cells that are dividing by mitosis producing daughter cells that are not yet fully differentiated
Initiation of Cancer at Cellular Level / Progression to Cancer
- What probably happens is: A single cell – perhaps an adult stem cell or progenitor cell in a tissue suffers a mutation (red line) in a gene involved in the cell cycle, e.g., an oncogene or tumor suppressor gene.
- This results in giving that cell a slight growth advantage over other dividing cells in the tissue.
- As that cell develops into a clone, some if its descendants suffer another mutation (red line) in another cell-cycle gene.
- This further deregulates the cell cycle of that cell and its descendants.
- As the rate of mitosis in that clone increases, the chances of further DNA damage increases.
- Eventually, so many mutations have occurred that the growth of that clone becomes completely unregulated.
- The result: full-blown cancer. Stem cells are cells that divide by mitosis to form either two stem cells, thus increasing the size of the stem cell “pool”, or one daughter that goes on to differentiate, and one daughter that retains its stem-cell properties.
- There is growing evidence that most of the cells in leukemias, breast, brain, and colon cancers are not able to proliferate out-of-control (and to metastasize).
- Only those members of the clone that retain their stem-cell-like properties (~2.5% of the cells in a tumor of the colon) can do so.
- There is certain logic to this.
- Most terminally-differentiated cells have limited potential to divide by mitosis and, seldom passing through S phase of the cell cycle, are limited in their ability to accumulate the new mutations that predispose to becoming cancerous.
- Furthermore, they often have short life spans – being eliminated by apoptosis (e.g., lymphocytes) or being shed from the tissue (e.g., epithelial cells of the colon).
- The adult stem cell pool, in contrast, is long-lived, and its members have many opportunities to acquire new mutations as they produce differentiating daughters as well as daughters that maintain the stem cell pool.
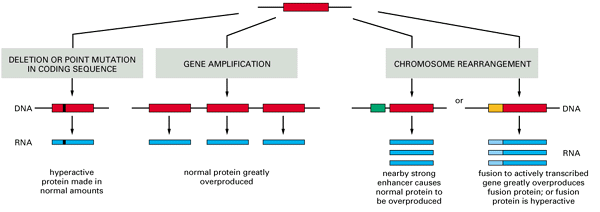
- Different types of genetic accident that can convert a proto-oncogene into an oncogene are summarized in Figure 3.49.
- The gene may be altered by a point mutation, by a deletion, through a chromosomal translocation, or by insertion of a mobile genetic element such as retroviral DNA.

Related posts:
 CYTOGENETICS OF ANEUPLOIDS AND STRUCTURAL HETEROZYGOTES
CYTOGENETICS OF ANEUPLOIDS AND STRUCTURAL HETEROZYGOTES
 Molecular Basis of Gene Mutation
Molecular Basis of Gene Mutation
 Concept of Cellular differentiation totipotency
Concept of Cellular differentiation totipotency
 Sex Linked Inheritance
Sex Linked Inheritance
 Chemical and Physical Mutagens
Chemical and Physical Mutagens
 Proto-oncogenes & Oncogenes
Proto-oncogenes & Oncogenes
 Inherited Human Disease Related with the Gene
Inherited Human Disease Related with the Gene
 Role of Induced Mutation in Crop Improvement
Role of Induced Mutation in Crop Improvement
 Sex Determination
Sex Determination

