Multiple Alleles
- The word allele is a general term to denote the alternative forms of a gene or contrasting gene pair that denote the alternative form of a gene is called allele.
- These alleles were previously considered by Bateson as hypothetical partner in Mendelian segregation.
- In Mendelian inheritance a given locus of chromosome was occupied by 2 kinds of genes, i.e., a normal gene (for round seed shape) and other its mutant recessive gene (wrinkled seed shape).
- But it may be possible that normal gene may show still many mutations in pea besides the one for wrinkledness.
- Here the locus will be occupied by normal allele and its two or more mutant genes.
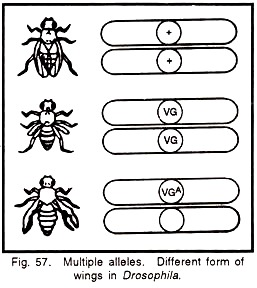
- Thus, three or more kinds of genes occupying the same locus in individual chromosome are referred to as multiple alleles.
- In short many alleles of a single gene are called multiple alleles.
- The concept of multiple alleles is described under the term “multiple allelism”.Dawson and Whitehouse in England proposed the term panallele for all the gene mutations at a given locus in a chromosome.
- These differ from the multiple factor in one respect that multiple factors occupy different loci while alleles occupy same locus.
- “Three or more kinds of gene which occupy the same locus are referred to as multiple alleles.” Altenburg
Characteristics of Multiple Alleles:
1. The study of multiple alleles may be done in population.
2. Multiple alleles are situated on homologous chromosomes at the same locus.
3. There is no crossing over between the members of multiple alleles. Crossing over takes place between two different genes only (inter-generic recombination) and does not occur within a gene (intragenic recombination).
4. Multiple alleles influence one or the same character only.
5. Multiple alleles never show complementation with each other. By complementation test the allelic and non-allelic genes may be differentiated well. The production of wild type phenotype in a trans-heterozygote for 2 mutant alleles is known as complementation test.
6. The wild type (normal) allele is nearly always dominant while the other mutant alleles in the series may show dominance or there may be an intermediate phenotypic effect.
7. When any two of the multiple alleles are crossed, the phenotype is of a mutant type and not the wild type.
8. Further, F2 generations from such crosses show typical monohybrid ratio for the concerned character.
Examples of Multiple Alleles:
1. Wings of Drosophila:
- In Drosophila wings are normally long.
- There occurred two mutations at the same locus in different flies, one causing vestigial (reduced) wings and other mutation causing antlered (less developed) wings.
- Both vestigial and antlered are alleles of the same normal gene and also of each other and are recessive to the normal gene.
- Suppose vestigial is represented by the symbol ‘vg’ and antlered wing by ‘vga‘. The normal allele is represented by the symbol +.
- Thus, there are three races of Drosophila:
(i) Long ++ (+/+)
(ii) Vestigial vg vg (vg/vg)
(iii) Antlered vga vga (vga/vga)
- A cross between a long winged normal fly and another having vestigial wings or antlered wings is represented below:
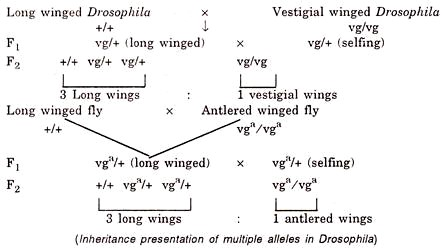
- When a fly with vestigial wing is crossed with another fly having antlered wings, the F1 hybrids are intermediate in wing length showing that none of the mutated gene is dominant over the other.
- This hybrid is some times said as the vestigial antlered compound and contains two mutated genes at the same locus.
- They show Mendelian segregation and recombination.
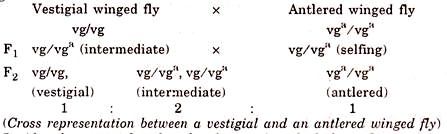
- Besides the vestigial and antlered wing described above there are several other mutations occurring at the same locus and resulting in nicked wings, strap wings or no wings etc.
- These are all multiple alleles.
Close Linkage Versus Allelism:
- If we assume that these mutant genes, vestigial and antlered are not allelic located at different loci in place of locating at same locus in different chromosomes so closely linked that there is no crossing over between them, the mutant gene will suppress the expression of adjacent normal allele to certain extent.
- These closely linked genes are called pseudo alleles and this suppression is the result of position effect.
- Thus, visible or apparent cases of allelism may be explained on the assumption of close linkage.
- Another example of multiple alleles is the eye colour in Drosophila.
- The normal colour of the eye is red.
- Mutation changed this red eye colour to white.
- Other mutations at white locus took place changing the red eye colour to various lighter shades like cherry, apricot, eosin, creamy, ivory, blood etc., are also visible and are due to multiple alleles.
- A cross between the two mutant forms, produces intermediate type in the F1 except white and apricot races which are not alleles but closely linked genes.
2. Coat Colour in Rabbit:
- The colour of the skin in rabbits is influenced by a series of multiple alleles.
- The normal colour of the skin is brown.
- Besides it there are white races called albino and Himalayan as the mutant races.
- The Himalayan is similar to albino but has darker nose, ear, feet and tail.
- The mutant genes albino (a) and Himalayan (ah) occupy the same locus and are allelic. Both albino and Himalayan are recessive to their normal allele (+).
- A cross between an albino and Himalayan produces a Himalayan in the F1 and not intermediate as is usual in the case of other multiple alleles.

3. Self-Sterility in Plants:
- Kolreuter (1764) described self- sterility in tobacco (Nicotiana longiflora).
- The reason was done by East.
- He described that self-sterility is due to series of alleles designated as s1, s2, s3 and s4 etc.
- The hybrids S1/S2 or S1/S3 or S3/S4 are self-sterile because pollen grains from these varieties did not develop, but pollens of S1/S2 were effective and capable of fertilization with S3/S4.
- The genes causing self-sterility in plants probably produce their effects by controlling the growth rate of the pollen tubes.
- In compatible combinations, the pollen tube grows more and more rapidly as it approaches the ovule, but in non-suitable ones, the growth of the pollen tube slows down considerably, so that the flower withers away before fertilization can take place.
4. Blood Groups in Man:
- Several genes in man produce multiple allelic series which affect an interesting and important physiological characteristic of the human red blood cells.
- The red blood cells have special antigens properties by which they respond to certain specific components (antibodies) of the blood serum.
- The antigen-antibody relationship is one of the great specificity like that between lock and key.
- Each antigen and its associated antibody has a peculiear chemical configuration.
- Landsteiner discovered in 1900 that when the red cells of one person are placed in the blood serum of another person, the cells become clumped or agglutinated.
- If blood transfusions were made between persons of two such incompatible blood groups, the transfused cells were likely to clump and shut out the capillaries in the recipient, some times resulting in death.
- However, such reactions occurred only when the cells of certain individuals were placed in serum from certain other persons.
- It was found that all persons could be classified in to four groups with regard to the antigen property of the blood cells.
- Large number of persons have been classified in to these four groups by means of the agglutination test and the distribution of blood groups in the offspring of parents of known blood groups has been studied.
- The evidence shows that these blood properties are determined by a series of three allelic genes IA, IB and i, as follows:

- IA is a gene for the production of the anti-gin A. IB for antigen B, and i for neither antigen.
- The existence of these alleles in man and the case with which the blood groups can be identified have obvious practical applications in blood transfusion, cases of disputed percentage and description of human populations.
- The alleles of these genes which affect a variety of biochemical properties of the blood, act in such a way that in the heterozygous compound IAIB, each allele exhibits its own characteristics and specific effect.
- The cells of the heterozygote contain both antigens A and B. On the other hand, IA and IB both show complete dominance over i, which lacks both antigens.
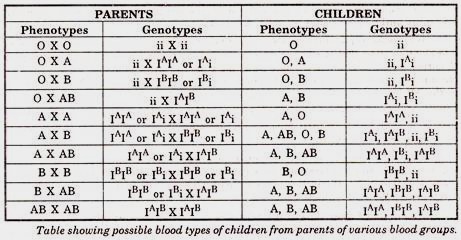
Table showing possible blood types of children from parents of various blood groups.
5. The ‘Rhesus’ Blood Group in Man:
- A very interesting series of alleles affecting the antigens of human blood has been discovered through the work of Landsteiner, Wiener, Race, Levine, Sanger, Mourant & several others.
- The original discovery was that the red cells are agglutinated by a serum prepared by immunizing rabbits against the blood of Rhesus monkey.
- The antigen responsible for this reaction was consequently called as Rhesus factor and the gene that causes this property was denoted as R-r or Rh-rh.
- Interest in this factor was stimulated by Levine’s study of a characteristic form of anaemia, known as Erythroblastosis foetalis, which occurs occasionally in new born infants.
- It was found that the infants suffering from this anaemia are usually Rh-positive and so are their fathers; but their mothers are Rh-negative.
- The origin of the disease was explained as follows:
- The Rh+ foetus developing in the uterus of an Rh– mother causes the formation of mother’s blood stream of anti Rh antibodies.
- These antibodies, especially as a result of a succession of several Rh+ pregnancies, gain sufficient strength in the mother’s blood so that they may attack the red blood cells of the foetus.
- The reaction between these antibodies of the mother and the red cells of her unborn child provokes haemolysis and anaemia; this may be serious enough to cause the death of the newborn infant or abortion of the foetus.
- The blood stream of a mother who has had an erythroblastotic infant is a much more potent and convenient reagent than sera of rabbits, immunized by blood of rhesus monkey’s for testing the blood of other persons to distinguish Rh+ from Rh– individuals using such sera from woman who had erythroblastotic infants, it was discovered that: there exist not one but several kinds of Rh+ and Rh- persons.
- There are several different Rh antigens which are detected by specific antisera.
- Thus, an Rh– woman immunized during pregnancy by the Rh+ children may have in her blood serum antibodies, that agglutinate not only Rh+ red cells but also cells from a few persons known to be Rh–.
- By selective absorption two kinds of antibodies may be separated from such a serum, one known as anti-D which agglutinates (= coagulates) only Rh+ cells, the other known as anti-C which agglutinates particular rare types of Rh–. Another specific antibody, known as anti-c agglutinates all cells that lack C.
- With these three antisera, six types of blood can be recognized.
- Studies of parent and children show that persons of type Cc are heterozygous for an allele C determining C anti-gena. CC persons are homozygous for C and cc are homozygous for c.
- There is obviously no dominance, each allele producing its own antigen in the heterozygote as in the AB blood type.
- No anti serum is available for detecting d, the alternative to D. D+ persons may be heterozygous or homozygous. However, the genotypes of such persons may be diagnosis from their progeny; for example D+ person who has a d– child is thereby shown to be Dd.
- Two other specific antibodies, anti-E and anti-c have been found. These detect the antigens E and e determined by a pair of alleles E and e.
- The three elementary types of antigens C-c, D and E-e, occur in fixed combinations that are always inherited together as alleles of a single gene.
- Wiener and Fisher showed the existence of a series of eight different alternative arrangements of these three types of Rh antigens and expressed them by means of following symbols.
The Rh System of Alleles:
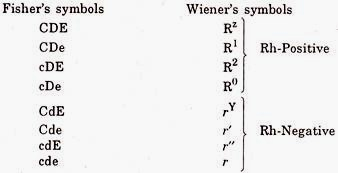
- Thus, allelism is determined by cross-breeding experiments.
- If one gene behaves as dominant to another the conclusion is that they are alleles and that they occupy identical loci in homologous chromosomes when two genes behave as dominant to other gene.
- They should occupy identical loci in the chromosome. When more than a pair of alleles occur in respect of any character in inheritance the phenomenon is known as multiple allelism.
- There is not much difference between the two theories of Wiener and Fisher.
- Wiener opinion is that there are multiple variations of one gene whereas according to the view of Fisher three different genes lying very close together are responsible for differences.
Pleiotropism:
- The opposite of polygene effect is known as pleiotropism i.e., a single gene influence or govern many characters.
- For example, gene for vestigial wing influence the nature of halters (modified balancers of Drosophila).
- The halters are not normal but reduced in flies with vestigial wings.
- The vestigial gene also affects position of dorsal bristles which instead of being horizontal turn out to be vertical.
- This gene also affects the shape of spermatheca i.e., the shape of spermatheca is changed ; the number of egg strings in the ovaries is decreased compared to normal when the vestigial larvae are well fed but relatively increased when they are poorly fed ; length of life and fruitfulness or fertility are lowered, and there are still other differences.
Theories of Allelism:
Various theories have been put forward to explain the nature of allelism origin and occurrence.
1. Theory of Point Mutation:
- According to this theory multiple alleles have developed as a result of mutations occurring at same locus but in different directions.
- Hence all the different wing lengths of Drosophila are necessarily the result of mutations which have occurred at same long normal wing locus in different directions.
2. Theory of Close Linkage or Positional Pseudoallelism:
- According to this view the multiple alleles are not the gene mutations at same locus but they occupy different loci closely situated in the chromosome.
- These genes closely linked at different loci are said to as pseudo alleles and affect the expression of their normal genes i.e., position effect.
3. Heterochromatin Theory of Allelism:
- Occasionally heterochromatin becomes associated with the genes as a result of chromosomal breakage and rearrangement.
- These heterochromatin particles suppress the nature of genes in question due to position effect.
- In maize the position effect are some times due to transposition (act of changing place or order) of very minute particles of heterochromatin.
- There are also sign or token that particles of different kinds of heterochromatin suppress the expression of normal gene to different degrees.
- In Drosophila the apricot might be a partially suppressed red (normal) and white completely suppressed red while apricot and white hybrid may give rise to red or intermediate by unequal crossing over.
- The above theories in some way or other do not explain clearly the particular case of allelism and it is possible that all the three theories are applicable in different cases.
Importance of Multiple Allelism:
- The study of multiple alleles has increased our knowledge of heredity.
- According to T.H. Morgan a great knowledge of the nature of gene has come from multiple alleles.
- These alleles suggest that a gene can mutate in different ways causing different effects.
- Multiple allelism also put forward the idea that different amounts of heterochromatin prevent the genes to different degree or space.
1. Pseudo alleles:
- Alleles are different forms of the same gene located at the corresponding loci or the same locus.
- Sometimes it has been found that non-homologous genes which are situated at near but different loci affect the same character in the same manner as if they are different forms or alleles of the same gene.
- They are said as pseudo alleles.
- These pseudo alleles which are closely linked show re-combinations by crossing over unlike the alleles.
2. Penetrance and Expressivity:
- Simply a recessive gene produces its phenotypic effect in homozygous condition and a dominant gene produces its phenotypic effect whether in homozygous or heterozygous condition.
- Some genes fail to produce their phenotypic effect when they should. The ability of a gene to produce its effect is called penetrance.
- The percentage of penetrance may be altered by changing the environmental conditions such as moisture, light intensity, temperature etc.
- A gene that always produces the expected effect is said to have 100 percent penetrance.
- If its phenotypic effect is produced only 60 percent of the individuals that contains it then it is said to show 60 percent penetrance.
- In Gossypium a mutant gene produces crinkled leaf.
- While all the leaves produced in the normal season are crinkled but some of the leaves
- which are produced late in the season do not show this character and are normal.
- It represents that penetrance is zero or in other words the gene is non-penetrant. Sometimes there is great variation in the manner in which a character is expressed in different plants.
- In Lima beans there is a variety named venturra where a dominant gene is responsible for tips and margins of the leaves of the seedlings to be partially deficient in chlorophyll. Sometimes only the margins are effected and sometimes only the tips.
- In other words, this single gene may express itself in a variety of ways that may resemble a number of characters.
- This gene is then to exhibit variable expressivity.
- Whether a gene is expressed at all is denoted by the term penetrance whereas the term expressivity denotes the degree of its expression.
3. Lsoalleles:
- Sometimes, a dominant gene occurs in two or more forms.
- These multiple dominant alleles will produce the same phenotypic effect in homozygous condition but their effect will show a small difference in heterozygous state.
- In Drosophila, thus, the gene for red eye colour is dominant over white.
- The red gene will produce dark red colour in the homozygous condition but in combination with the white allele the gene for red colour produces a dark red colour in flies from Soviet Russia but the same combination in the flies coming from the U.S.A. produces a light red colour.
- It does mean that dominant gene for red colour occurs in two forms.
- These are said as isoalleles.
4. Phenocopy:
- Characters are the result of interaction between the genotype and the environment. When a gene mutates, its phenotypic effect also changes.
- Some times, a change in the environment produces a visible change in the phenotype of the normal gene which resembles the effect as already known mutant.
- The effect of the normal gene under the changed environment is a mimic or imitation of the mutant gene. Such an imitation induced by environmental changes has been termed as phenocopy by Goldschmidt.
- In fowls, a mutant gene is responsible for the character, ruinplessness, in which the caudal vertebrate and tail feathers do not develop.
- Rumplessness is also induced as a phenocopy when normal eggs which do not have the gene for rumplessness, are treated with insulin before incubation.
- Phenocopies of other mutant genes are also produced in Drosophila by high temperature treatment of the larvae for short periods. It has also been found that different or non- allelic genes can produce the same phenotype.
- This phenomenon is said as genetic mimic or genocopy.
5. Xenia and Metaxenia:
- The immediate effect of foreign pollen on visible characters of the endosperm is called xenia.
- The ‘xenia’ term was given by Focke (1800).
- This has been studied in maize plant. If a white endosperm variety is open pollinated in the field where there are also plants of the yellow endosperm variety then the cobs that develop will contain a mixture of yellow and white seeds.
- The yellow colour of the endosperm in the yellow seeds is the result of fertilization by pollen from the yellow variety.
- The yellow colour indicates that the seeds are hybrids and the white seeds are homozygous.
- The yellow colour of the endosperm is dominant over white and when the plants raised from the yellow seeds are self-pollinated, yellow and white seeds are produced in the ratio of 3:1.
- Another example of xenia may be exemplified.
- If a sweet corn (maize) is pollinated by a starchy variety, the endosperm is starchy because the starchy gene introduced by the pollen is dominant over its sugary allele.
6. Metaxenia:
- It is the term used to describe the effect of foreign pollen on other tissues belonging to the mother plant, outside the endosperm and embryo.
- It is sometimes evident in the fruit and seed coats.
- In cucurbitaceous fruits, the skin colour is affected by the pollen grains; in oranges, the colour and flavour of the fruit is influenced by the pollen parent.
- The same is true of fuzziness and hair length in cotton.
- It has been suggested that metaxenia effects may be due to certain hormones secreted by the endosperm and embryo.

Related posts:
 Role of Induced Mutation in Crop Improvement
Role of Induced Mutation in Crop Improvement
 Molecular Basis of Gene Mutation
Molecular Basis of Gene Mutation
 CYTOGENETICS OF ANEUPLOIDS AND STRUCTURAL HETEROZYGOTES
CYTOGENETICS OF ANEUPLOIDS AND STRUCTURAL HETEROZYGOTES
 Proto-oncogenes & Oncogenes
Proto-oncogenes & Oncogenes
 Sex Linked Inheritance
Sex Linked Inheritance
 EUPLOIDY
EUPLOIDY
 Sex Determination
Sex Determination
 MUTATIONS -Spontaneous & Induced Mutations
MUTATIONS -Spontaneous & Induced Mutations

