![]()
Cell cycle and apoptosis
- The cell cycle, or cell-division cycle, is the series of events that take place in a eukaryotic cell leading to its replication.
- These events can be divided in two brief periods: interphase during which the cell grows, accumulating nutrients needed for mitosis and duplicating its DNA and the mitotic (M) phase, during which the cell splits itself into two distinct cells, often called “daughter cells”.
- The cell-division cycle is a vital process by which a single-celled fertilized egg develops into a mature organism, as well as the process by which hair, skin, blood cells, and some internal organs are renewed.
Phases of the cell cycle
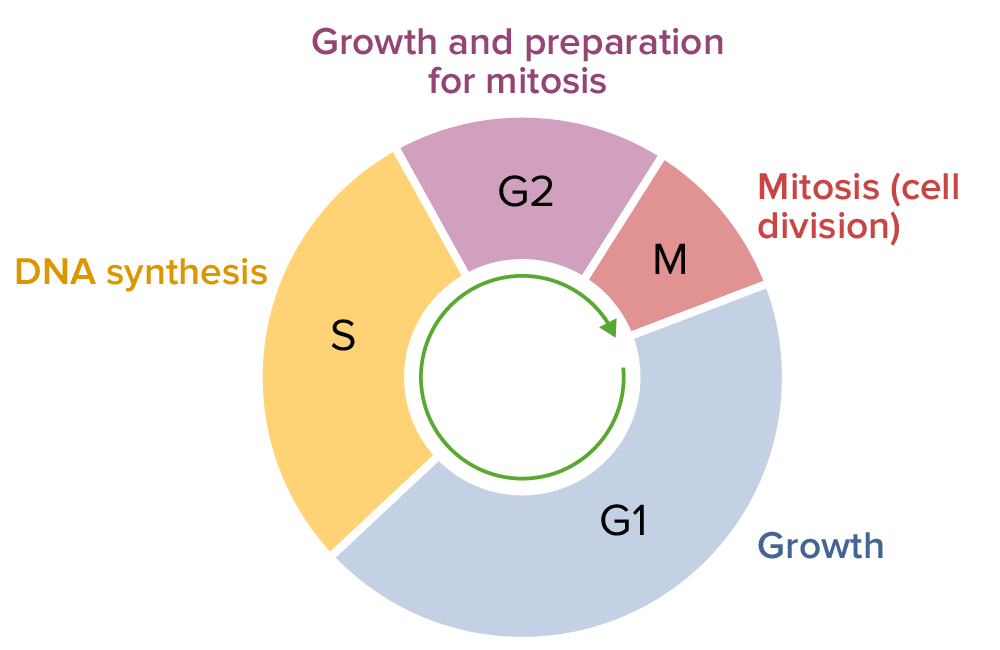
- The cell cycle consists of four distinct phases: G1 phase, S phase, G2 phase (collectively known as interphase) and M phase.
- M phase is itself composed of two tightly coupled processes: mitosis, in which the cell’s chromosomes are divided between the two daughter cells, and cytokinesis, in which the cell’s cytoplasm divides forming distinct cells.
- Activation of each phase is dependent on the proper progression and completion of the previous one.
- Cells that have temporarily or reversibly stopped dividing are said to have entered a state of quiescence called G0 phase.
M phase
- The relatively brief M phase consists of nuclear division (karyokinesis) and cytoplasmic division (cytokinesis).
- In plants and algae, cytokinesis is accompanied by the formation of a new cell wall.
- The M phase has been broken down into several distinct phases, sequentially known as prophase, Prometaphase, metaphase, anaphase and telophase leading to cytokinesis.
Interphase
- After M phase, the daughter cells each begin interphase of a new cycle.
- Although the various stages of interphase are not usually morphologically distinguishable, each phase of the cell cycle has a distinct set of specialized biochemical processes that prepare the cell for initiation of cell division.
G1 phase
- The first phase within interphase, from the end of the previous M phase till the beginning of DNA synthesis is called G1 (G indicating gap or growth).
- During this phase the biosynthetic activities of the cell, which had been considerably slowed down during M phase, resume at a high rate.
- This phase is marked by synthesis of various enzymes that are required in S phase, mainly those needed for DNA replication.
- Duration of G1 is highly variable, even among different cells of the same species.
S phase
- The S phase starts when DNA synthesis commences; when it is complete, all of the chromosomes have been replicated, i.e., each chromosome has two (sister) chromatids.
- Thus, during this phase, the amount of DNA in the cell has effectively doubled, though the ploidy of the cell remains the same.
- Rates of RNA transcription and protein synthesis are very low during this phase.
- An exception to this is histone production, most of which occurs during the S phase.
- The duration of S phase is relatively constant among cells of the same species.
G2 phase
- The cell then enters the G2 phase, which lasts until the cell enters mitosis.
- Again, significant protein synthesis occurs during this phase, mainly involving the production of microtubules, which are required during the process of mitosis.
- Inhibition of protein synthesis during G2 phase prevents the cell from undergoing mitosis.
G0 phase
- The term “post-mitotic” is sometimes used to refer to both quiescent and senescent cells.
- Non-proliferative cells in multicellular eukaryotes generally enter the quiescent G0 state from G1 and may remain quiescent for long periods of time, possibly indefinitely (as is often the case for neurons).
- This is very common for cells that are fully differentiated.
- Cellular senescence is a state that occurs in response to DNA damage or degradation that would make a cell’s progeny nonviable; it is often a biochemical alternative to the self-destruction of such a damaged cell by apoptosis.
Cell cycle inhibitors
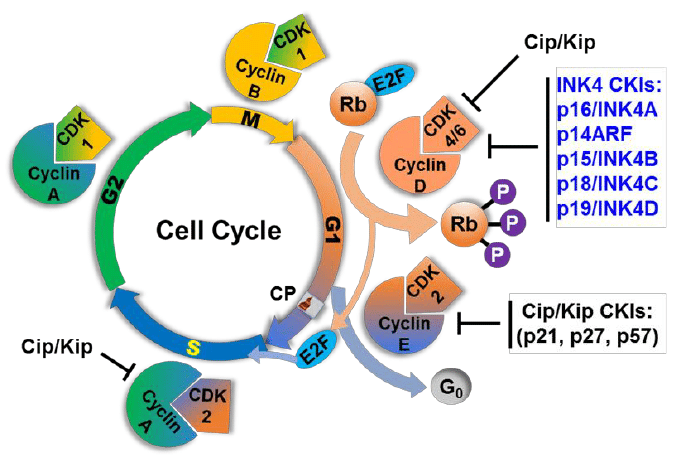
- Two families of genes, the cip/kip family and the INK4a/ARF (Inhibitor of Kinase 4/Alternative Reading Frame) prevent the progression of the cell cycle.
- Because these genes are instrumental in prevention of tumor formation, they are known as tumor suppressors.
- The cip/kip family includes the genes p21, p27 and p57.
- They halt cell cycle in G1 phase, by binding to, and inactivating, cyclin-CDK complexes. p21 is activated by p53 (which in turn is triggered by DNA damage e.g. due to radiation). p27 is activated by
Transforming Growth Factor β (TGF β), a growth inhibitor.
The INK4a/ARF family includes p16INK4a, which binds to CDK4 and arrests the cell cycle in G1 phase, and p14arf which prevents p53 degradation. And the amount of chromosomes are able to double at the same rate as in phase 2
Checkpoints
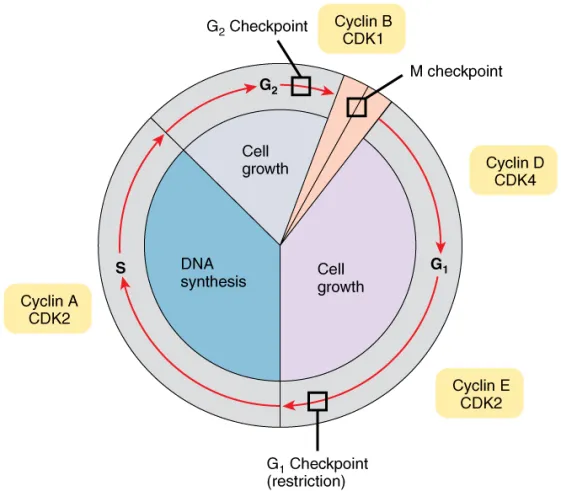
- Cell cycle checkpoints are used by the cell to monitor and regulate the progress of the cell cycle.
- Checkpoints prevent cell cycle progression at specific points, allowing verification of necessary phase processes and repair of DNA damage.
- The cell cannot proceed to the next phase until checkpoint requirements have been met.
- Several checkpoints are designed to ensure that damaged or incomplete DNA is not passed on to daughter cells.
- Two main checkpoints exist: the G1/S checkpoint and the G2/M checkpoint. G1/S transition is a rate-limiting step in the cell cycle and is also known as restriction point.
- An alternative model of the cell cycle response to DNA damage has also been proposed, known as the post replication checkpoint. p53 plays an important role in triggering the control mechanisms at both G1/S and G2/M checkpoints.
The development of the term apoptosis
- Already since the mid-nineteenth century, many observations have indicated that cell death plays a considerable role during physiological processes of multicellular organisms, particularly during embryogenesis and metamorphosis .
- The term programmed cell death was introduced in 1964, proposing that cell death during development is not of accidental nature but follows a sequence of controlled steps leading to locally and temporally defined self-destruction
- Eventually, the term apoptosis had been coined in order to describe the morphological processes leading to controlled cellular self-destruction and was first introduced by Kerr, Wyllie and Currie in 1972.
- Apoptosis is of Greek origin, having the meaning “falling off or dropping off”, in analogy to leaves falling off trees or petals dropping off flowers.
- This analogy emphasizes that the death of living matter is an integral and necessary part of the life cycle of organisms.
- The apoptotic mode of cell death is an active and defined process which plays an important role in the development of multicellular organisms and in the regulation and maintenance of the cell populations in tissues upon physiological and pathological conditions.
- It should be stressed that apoptosis is a well-defined and possibly the most frequent form of programmed cell death, but that other, non-apoptotic types of cell death also might be of biological significance .
The significance of apoptosis
- The development and maintenance of multicellular biological systems depends on a sophisticated interplay between the cells forming the organism, it sometimes even seems to involve an altruistic behaviour of individual cells in favour of the organism as a whole.
- During development many cells are produced in excess which eventually undergo programmed cell death and thereby contribute to sculpturing many organs and tissues.
- Taken together, apoptotic processes are of widespread biological significance, being involved in e.g. development, differentiation, proliferation/homoeostasis, regulation and function of the immune system and in the removal of defect and therefore harmful cells.
- Thus, dysfunction or deregulation of the apoptotic program is implicated in a variety of pathological conditions.
- Defects in apoptosis can result in cancer, autoimmune diseases and spreading of viral infections, while neurodegenerative disorders and AIDS diseases are caused or enhanced by excessive apoptosis.
- Due to its importance in such various biological processes, programmed cell death is a widespread phenomenon, occurring in all kinds of metazoans such as in mammals, insects, nematodes.
- Moreover, programmed cell death also might play a role in plant biology, and apoptosis-like cell death mechanisms even have been observed and used as a model system in yeast.
- Fascinating insights into the origin and evolution of programmed cell death might possibly be given by the fact, that programmed cell death is also an integral part of the life cycle of other unicellular eukaryotes and that even prokaryotes.
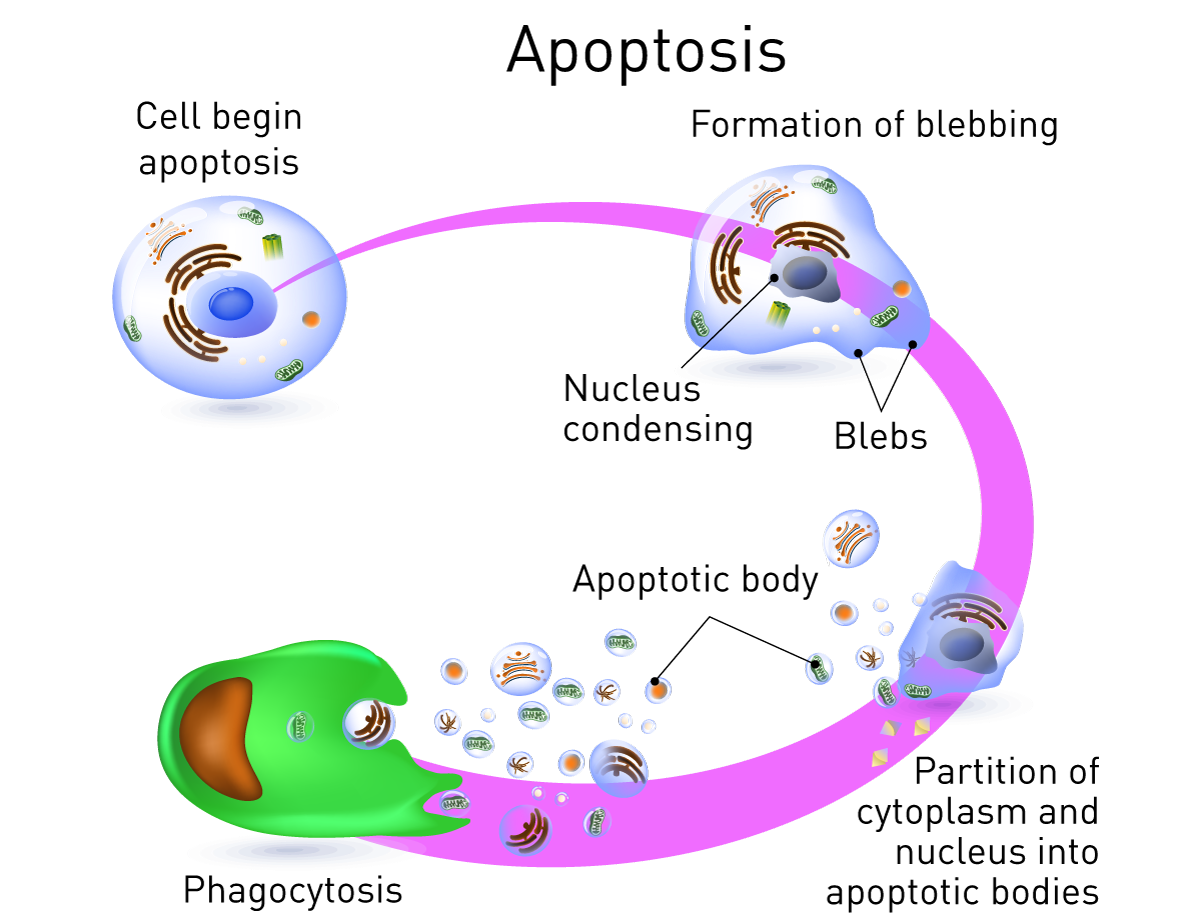
Morphological features of apoptosis
- Apoptotic cells can be recognized by stereotypical morphological changes: the cell shrinks, shows deformation and looses contact to its neighboring cells.
- Its chromatin condenses and migrate at the nuclear membrane and finally the cell is fragmented into compact membrane-enclosed structures, called ‘apoptotic bodies’ which contain cytosol, the condensed chromatin, and organelles.
- The apoptotic bodies are engulfed by macrophages and thus are removed from the tissue without causing an inflammatory response.
- Those morphological changes are a consequence of characteristic molecular and biochemical events occurring in an apoptotic cell, most notably the activation of proteolytic enzymes which eventually mediate the cleavage of DNA into oligo-nucleosomal fragments as well as the cleavage of a multitude of specific protein substrates which usually determine the integrity and shape of the cytoplasm or organelles .