The Structure Analysis of Eukaryotes Introns and their Significance
- Introns, derived from the term “intragenic regions”, are non-coding sections of precursor mRNA (pre-mRNA) or other RNAs, that are removed (spliced out of the RNA) before the mature RNA is formed.
- Once the introns have been spliced out of a pre-mRNA, the resulting mRNA sequence, composed of exons, is ready to be translated into a protein.
- The corresponding parts of a gene are known as introns as well.
Introduction –
- Introns are common in eukaryotic pre-mRNA, but in prokaryotes they are only found in tRNA and rRNA. Introns, which are non-coding sections of a gene that are removed, are the opposite of exons which remain in the mRNA sequence after processing.
- The number and length of introns varies widely among species, and among genes within the same species.
- Genes of higher organisms, such as mammals and flowering plants, have numerous introns, which can be much longer than the nearby exons.
- Some less advanced organisms, such as fungus Saccharomyces cerevisiae, and protists, have very few introns. In humans, the gene with the greatest number of introns is the gene for the protein Titin, with 362 introns.
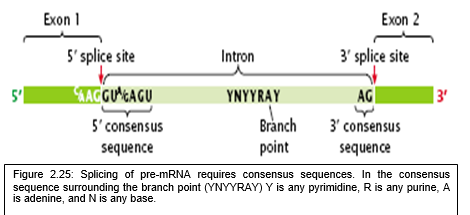
- Figure 2.25 is showing simple illustration of a pre-mRNA, with introns (top), after the introns have been removed via splicing, the mature mRNA sequence is ready for translation (bottom).
- Introns sometimes allow for alternative splicing of a gene, so that several different proteins which share some sequences in common can be translated from a single gene.
- The control of mRNA splicing is performed by a wide variety of signaling molecules.
- Introns may also contain “old code”, or sections of a gene that were once translated into a protein, but have since been discarded.
- It was generally assumed that the sequence of any given intron is junk DNA with no function. More recently, however, this is being disputed.
- Introns contain several short sequences that are important for efficient splicing.
- The exact mechanism for these intronic splicing enhancers is not well understood, but it is thought that they serve as binding sites on the transcript for proteins which stabilize the spliceosome.
- It is also possible that RNA secondary structure formed by intronic sequences may have an effect on splicing.
Discovery –
- The discovery of introns led to the Nobel Prize in Physiology or Medicine in 1993 for Phillip Allen Sharp and Richard J. Roberts.
- The term intron was introduced by American biochemist Walter Gilbert:
- “The notion of the cistron […] must be replaced by that of a transcription unit containing regions which will be lost from the mature messenger – which I suggest we call introns (for intragenic regions) – alternating with regions which will be expressed – exons.” (Gilbert 1978)
Classification of Introns –
- Some introns, such as Group I and Group II introns, are actually ribozymes that are capable of catalyzing their own splicing out of a primary RNA transcript.
- This self splicing activity was discovered by Thomas Cech, who shared the 1989 Nobel Prize in Chemistry with Sidney Altman for the discovery of the catalytic properties of RNA.
Four classes of introns are known to exist:
- Group I intron
- Group II intron
- Group III intron
- Nuclear introns
- Sometimes group III introns are also identified as group II introns, because of their similarity in structure and function.
- Nuclear or spliceosomal introns are spliced by the spliceosome and a series of snRNAs (small nuclear RNAs).
- There are certain splice signals (or consensus sequences) which abet the splicing (or identification) of these introns by the spliceosome.
- Group I, II and III introns are self splicing introns and are relatively rare compared to spliceosomal introns.
- Group II and III introns are similar and have a conserved secondary structure.
- The lariat pathway is used in their splicing.
- They perform functions similar to the spliceosome and may be evolutionarily related to it.
- Group I introns are the only class of introns whose splicing requires a free guanine nucleoside.
- They possess a secondary structure different from that of group II and III introns.
- Many self-splicing introns code for maturases that help with the splicing process, generally only the splicing of the intron that encodes it.
Intron evolution –
- There are two competing theories that offer alternative scenarios for the origin and early evolution of spliceosomal introns (Other classes of introns such as self-splicing and tRNA introns are not subject to much debate, but see for the former).
- These are popularly called as the Introns-Early (IE) or the Introns-Late (IL) views.
- The IE model, championed by Walter Gilbert, proposes that introns are extremely old and numerously present in the earliest ancestors of prokaryotes and eukaryotes (the progenote).
- In this model introns were subsequently lost from prokaryotic organisms, allowing them to attain growth efficiency.
- A central prediction of this theory is that the early introns were mediators that facilitated the recombination of exons that represented the protein domains.
- Such a model would directly lead to the evolution of new genes.
- Unfortunately, the model cannot account for the variations in the positions of shared introns between different species.
- The IL model proposes that introns were more recently inserted into original intron-less contiguous genes after the divergence of eukaryotes and prokaryotes.
- In this model, introns probably had their origin in parasitic transposable elements.
- This model is based on the observation that the spliceosomal introns are restricted to eukaryotes alone.
- However, there is considerable debate on the presence of introns in the early prokaryote-eukaryote ancestors and the subsequent intron loss-gain during eukaryotic evolution.
- It is also suggested that the evolution of introns and more generally the intron-exon structure is largely independent of the coding-sequence evolution.
Identification
- Nearly all eukaryotic nuclear introns begin with the nucleotide sequence GU, and end with AG (the GU-AG rule).
- These, along with a larger consensus sequence, help direct the splicing machinery to the proper intronic donor and acceptor sites.
- This mainly occurs in eukaryotic primary mRNA transcripts.



