![]()
Genetic conjugation in Bacteria
Conjugation:
- Lederberg and Tatum (1946) discovered conjugation in E. coli and its detailed studies were made by Woolman and Jacob (1956).
- Conjugation, is a process by which genetic material is transferred from one bacterial cell (“donor cell” or “male cell”) to another (“recipient cell” or “female cell”) through a specialized intercellular connection called sex-pilus or conjugation tube.
- The maleness and femaleness of bacterial cells are determined by the presence or absence of F-plasmid (also called F-factor or sex factor).
- F- plasmid, an extra chromosomal genetic material, is always present in the cytoplasm of donor or male cells, and the latter develop specialized cell surface appendages called F-pili or sex-pili under the control of F-plasmid.
- Recipient or female cells always lack F-pIasmids and, therefore, F-pili are not present on their surface.
- In this process, the exchange of genetic material takes place through a conjugation tube between the two cells of bacteria.
- The process was first postulated by Joshua Lederberg and Edward Tatum (1946) in Escherichia coli.
- They were awarded the Nobel Prize in 1958 for their work on bacterial genetics.
- Later on, it has also been demonstrated in Salmonella, Vibrio and Pseudomonas.
- There are two mating types of bacteria, one is. male type or F+ or donor cell,
- which donates some DNA.
- The other one is female type or F– or recipient cell, which receives DNA.
- Later, after receiving DNA, the recipient cell may behave as donor cell i.e., F+ type.
- The F-factor is the fertility factor, sex-factor or F-plasmid present in the cell of F+ i.e., donor cell or male type.
- The plasmid takes part in conjugation is called episome.
- In this process, two cells of opposite mating type i.e., F+ and F– become temporarily attached with each other by sex pilus .
- The sex pilus has a hole of 2.5 pm diameter through which DNA can pass from donor to recipient cell.
- The F-factor or F-plasmid is a double stranded DNA loop, present in the cytoplasm; apart from the nucleoid.
- The F-factor contains about 20 genes.
- After the establishment of conjugation tube, the F-factor prepares for replication by the rolling circular mechanism. he two strands of F- factor begin to separate from each other and one of them passes to the recipient i.e., F– cell.
- After reaching in F– cell, enzymes synthesise a complementary strand that forms a double helix, which bends into a loop.
- The conversion process is thus completed. In the donor cell i.e., in F+, a new DNA strand also forms to complement the left over DNA strand of the F-factor.
- There is another type of conjugation where passage of nucleoid DNA takes place through conjugation tube.
- Strains of bacteria are known as Hfr (high frequency of recombination) strain.
- William Hayes discovered such strains of E. coli in 1950s.
- The Hfr factor is also called episome. In Hfr strain, the F-factor is attached with the nucleoid DNA i.e., the bacterial chromosome.
- In this process, Hfr and F– cells become attached with each other by sex pilus (Fig. 2.27). At the point of attachment of F-factor, the bacterial chromosome opens and a copy of one strand is formed by the rolling circular mechanism.
- A portion of single stranded DNA then passes into the recipient cell through pilus. Due to agitation in medium, the conjugation tube may not survive for long time because of broken pilus.
- Thereby, the total length of transfer DNA may not be able to take entry to the recipient cell.
The behaviour of the transferred DNA depends on the presence and absence of F-factor:
- If F-factor is indeed transferred, then it usually remains detached from the chromosome of recipient cell and enzymes synthesise a complementary DNA strand.
- The factor then forms a loop and exists as a plasmid, thereby the recipient cell becomes a donor.
- If F-factor remains at the rear end of the transfer DNA during its entry to the recipient cell, the F-factor may not be able to take entry due to broken pilus and only a portion with new genes (Fig. 2.27) takes up the entry.
- Thereby, the F– strain remains as recipient one. In F– strain, genetic recombination takes place between donor fragment and recipient DNA.
- Sometimes, if the F-factor gets free from the Hfr cell and maintains an independent status, then the Hfr cell converts to a F+ cell. Sometimes during the leaving of F-factor from the bacterial chromosome, it takes a segment of chromosomal DNA.
- The F-factor with segment of chromosomal DNA is called F’-factor.
- Later on, during conjugation, when this F’-factor is transferred, the recipient cell receives some chromosomal DNA from the donor cell.
- This process is called sexduction. In this process, the recipient cell receives a portion of chromosomal DNA which duplicates with the existing one for a specific function, thereby the recipient cell is a partial diploid.
F-plasmid or F-factor can exist in two different states:
(i) The autonomous state in which it lies free in the cytoplasm and replicate independent of the bacterial chromosome (DNA); a donor or male cell containing F- factor in autonomous state is called F+ cell, and
(ii) The integrated state in which it is integrated (inserted) into the bacterial chromosome (DNA) and replicate along with it; a donor or male cell containing F-factor in integrated state is called Hfr cell (for high frequency recombination) or high frequency male cell. However, the recipient or female cell lacks F-factor and this is called F– cell.
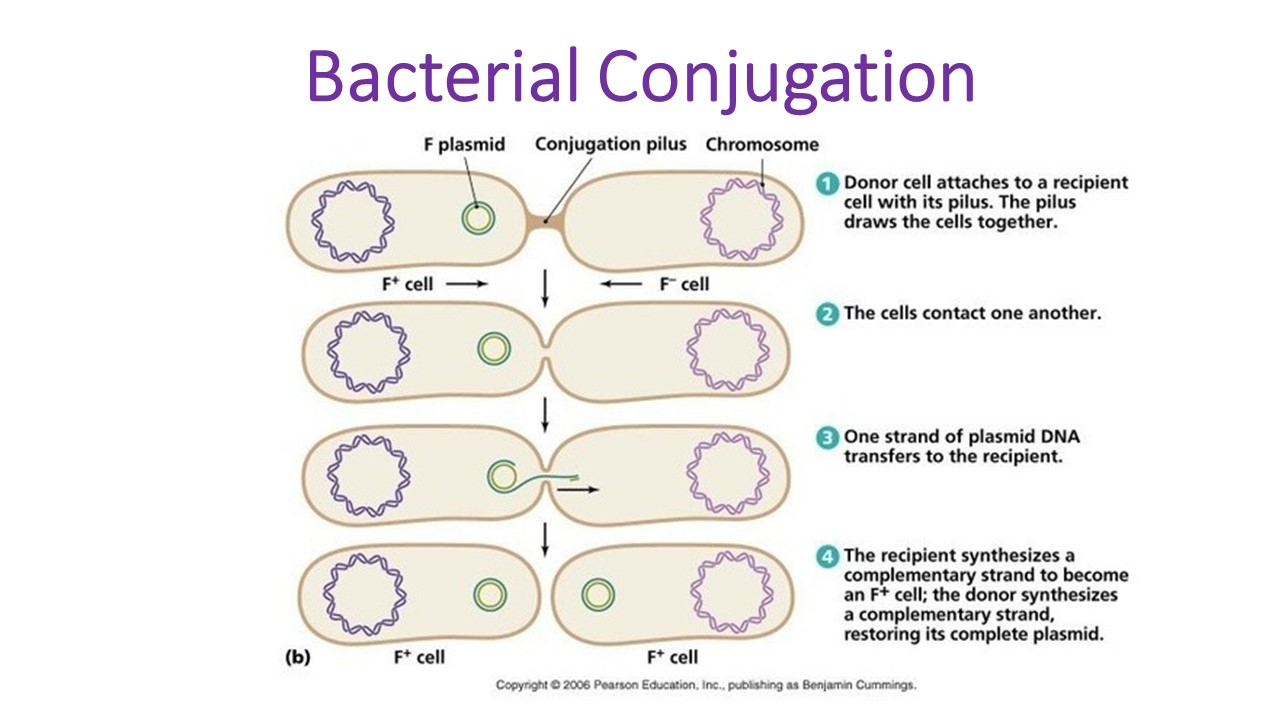
1. Conjugation between a F+ (donor) cell and a F– (recipient) cell:
- In conjugation between a F+ (donor) cell and a F– (recipient) cell, it is the autonomous F-factor (F-plasmid) which is transferred, never the bacterial DNA (Fig. 29.2).
- When the two cells (F+ and F–) come close to each other, the F-pilus of the F+ (donor) cell attaches with the F– (recipient) cell and acts as a conjugation tube.
- Simultaneously, the double-stranded circular F-factor DNA is nicked at a specific point, and begins to replicate producing a single-stranded copy of the F- factor DNA, which migrates through the tube into the cytoplasm of the F– (recipient) cell.
- It becomes double- stranded, and circulars and lies free in the cytoplasm thus rendering the recipient cell to become F+ donor cell. In this way, mixing a population of F+ (donor) cells with a population of F+ (recipient) cells results in the conversion of virtually all the cells in the population becoming F+ (donor) cells.

Diagrammatic representation of conjugation between F+ cell and F- cell
2. Conjugation between Hfr Donor Cells and Recipient (F–) Cell:
- The Hfr donor cells are considered to be fertile because, unlike F+ (donor) cells, their chromosomal segments are transferred from donor to recipient cells and the F-factor remains in situ.
- When the two cells (Hfr and F–) come in contact, a conjugation tube develops between them. The circular DNA of Hfr donor cell is nicked and replication is initiated.
- The integrate F-factor always lies at the rear end of the DNA molecule.
- The replication of DNA starts towards the end near the conjugation tube and the newly synthesized single strand starts migrating through the tube into the recipient (F–) cell.
- In nature, the mating of two cells exists for a short period and gets interrupted resulting in the migration of only a portion of the donor DNA into the recipient cell. Since the F-factor lies at the rear end of the molecule, it is rarely transferred to the recipient cell
- The genes of the newly entered DNA fragment may replace the homolgous genes of the DNA of the recipient cell, resulting in a recombinant genetic material.
- The newly formed recombinant genetic material now possesses those male characters that have been transferred through recombination with the migrated DNA fragment (Fig. 29.3).

Diagrammatic representation of cojugation between Hfr donor cell and F- cell
3. Conjugation between F’ (F-prime) Male and F– (Recipient) Cell (Sex-Duction):
- Existence of Hfr donor cells is not absolute.
- The F-factor integrated into the bacterial DNA of Hfr donor cells may dissociate and become free in the cytoplasm (Fig. 29.4).
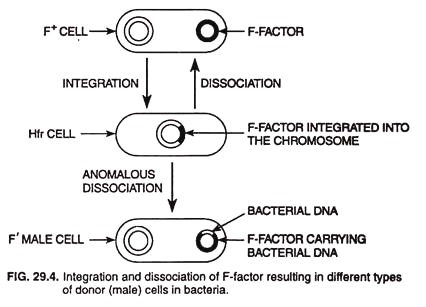
- The dissociation may be occasionally anomalous during which the dissociated F-factor may bring with it some genes of the bacterial chromosome.
- Adelberg and Burns (1958) first identified such a modified F-factor and called it F’ (“F-prime”) factor; the donor cell possessing this factor is called F’ (F-prime) male.
- When a F’ male conjugates with F– (recipient) cell, the F’-factor is transferred from donor to the recipient cell, and such a recipient bacterial cell becomes heterozygous (merozygous) for that part of the bacterial chromosome, which the F’- factor had obtained during its anomalous dissociation.
- Transfer of F’-factor to recipient cell apparently occurs by the same mechanism as F-factor, transfers during in F+ and F– mating and chromosome transfer in Hfr and F– cell mating.
- Genetic recombination of this type, mediated by F’-factor, is called sex-duction or F-duction.
- Integration and dissociation of F-factor resulting in diffrent types of donor cells in bacteria



