![]()
Structure and function of microbodies
Microbodies : Structure and Types
- Microbodies are spherical or oblate in form.
- They are bounded by a single membrane and have an interior or matrix which is amorphous or granular Micrbodies are most easily distinguished from other cell organelles by their content of catalase enzyme.
- Catalase can be visualized with the electron microscope when cells are treated with the stain DAB (i.e., 3,3′-diaminobenzidine).
- The product is electron opaque and appears as dark regions in the cell where catalase is present
- The technique of isolation of microbodies of plant tissues includes the following steps :
- (1) Tissues are group very carefully to save microbodies from disruption.
- (2) The homogenate is with differential centrifugation to obtain a fraction of the cell homogenate which is rich in microbodies.
- (3) The enriched fraction is subjected to isopycnic ultra-centrifugation on discontinuous or continuous sucrose density gradient.
- Recent biochemical studies have distinguished two types of microbodies, namely peroxisomes and glycoxysomes.
- These two organelles differ both in their enzyme complement and in the type of tissue in which they are found.
- Peroxisomes are found in animal cells and the leaves of higher plants.
- They contain catalases and oxidases (e.g., D-amino oxidase and urate oxidase).
- In both they participate in the oxidation of substrates, producing hydrogen peroxide which is subsequently destroyed by catalase activity.
- In plant cells, peroxisomes remain associated with ER. chloroplasts and mitochondria and are involved in photorespiration.
- Gloxysomes occur only in plant cells and are particularly abundant in germinating seeds which store fats as a reserve food material.
- They contain enzymes of glyoxylate cycle besides the catalases and oxidases
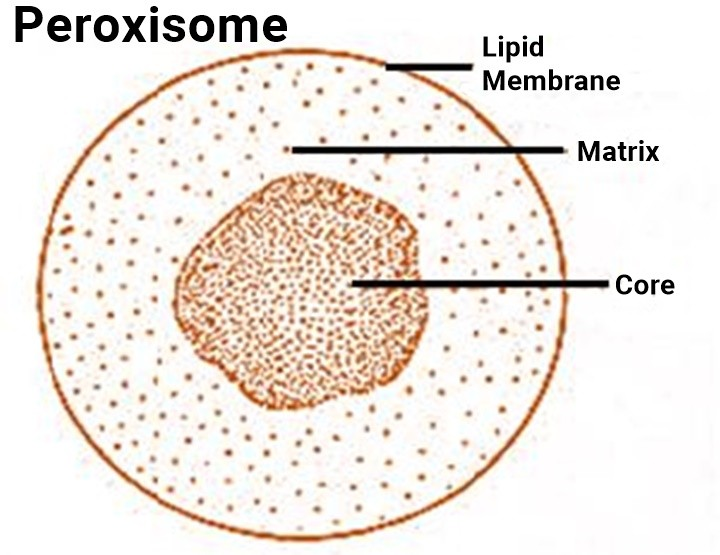
Peroxisomes
- Peroxisomes occur in many animal cells and in a wide range of plants.
- They are present in all photosynthetic cells of higher plants in etiolated leaf tissue,
- in coleoptiles and hypocotyls, in tobacco stem and callus, in ripening pear fruits and also in Euglenophyta, Protozoa, brown algae, fungi liverworts, mosses and ferns.
- Peroxisomes are variable in size and shape, but usually appear circular in cross section having diameter between 0.2 and 1.5mm (0.2 and 0.25 mm diameter in most mammalian tissues : 0.5mm diameter in rat liver cells).
- They have a single limiting unit membrane of lipid and protein molecules, which encloses their granular matrix. In some cases (e.g., in the festuciod grasses) the matrix contains numerous threads or fibrils, while in others they are observed to contain either an amorphous nucleoid or a dense inner core which in many species shows a regular crystalloid structure (e.g., tobacco leaf cell).
- Little is known about the function of the core, except that it is the site of the enzyme urate oxidase in rat liver peroxisomes and much of the catalase in some plants .
- Recently, a possible relationship has been stressed between peroxides and free radicals (such as superoxide anion -O2–) with the process of aging.
- These radicals may act on DNA molecule to produce mutations altering the transcription into mRNA and the translation into proteins.
- In addition, free radicals and peroxides can affect the membranes by causing peroxidation of lipids and proteins.
- For these reasons reducing compounds such as vitamin E or enzymes such as superoxide dismutase could play a role in keeping the healthy state of a cell.
Glycolate cycle-
- Peroxisomes of plant leaves contain catalaze together with the enzymes of glycolate pathway, as glycolate oxidase, glutamate glyoxylate, serine-glyoxylate and aspirate-a-ketoglutarate aminotransferases, hydroxyl pyruvate reductase and malic dehydrogenase.
- They also contain FAD, NAD and NADP coenzymes.
- The glycolate cycle is thought to bring about the formation of the amino acids-glycine and serine-from the non-phosphorylated intermediates of photosynthetic carbon reduction cycle, i.e., glycerate to serine, or glycolate to glycine and serine in a sequence of reactions which involve chloroplasts, peroxisomes, mitochondria and cytosol.
- The glycolate pathway also generates C1 compounds and serves as the generator of precursors for nucleic acid biosynthesis.

Photorespiration–
- In green leaves, there are peroxisomes that carry out a process called photorespiration which is a light – stimulated production of CO2 that is different from the generation of CO2 by mitochondria in the dark.
- In photorespiration, glycolic acid (glycolate), a two-carbon product of photosynthesis is released from chloroplasts and oxidized into and H2O2 by a peroxisomal enzyme called glycolic acid oxidase.
- Later on, glyoxylate is oxidized into CO2 and formate:
CH2OH.COOH + O2 ® CHO – COOH + H2O2
CHO – COOH + H2O2 ® HCOOH + CO2 + H2O
- Photorepiration is so called because light induces the synthesis of glycolic acid in chloroplasts.
- This process involves intervention of two organelles : cloroplasts and peroxisomes.
- Lastly, photorespiration is driven by atmospheric conditions in which the O2 tension is high and CO2 tension low.
- Apparently O2 competes with CO2 for the enzyme ribulose diphosphate carboxylase which normally is the key enzyme in CO2 fixation during photosynthesis.
- When O2 is used by the enzyme, an unstable intermediate is formed which breaks down into 3-phosphoglycerate and phosphoglycolate.
- The latter tends to increase the glycolate concentration by removal of its phosphate group and, therefore, more glycolate is available for additional oxidation and CO2 release
- Photorespiration is a wasteful process for the plant cell, since, it significantly reduces the efficiency of the process of photosynthesis (i.e., it returns a portion of fixed CO2 to the atmosphere).
- It is a particular problem in C3 plants that are more readily affected by low CO2 tensions ; C4 plants are much more efficient in this regard.

Biogenesis of Peroxisomes
- At one time it was thought that the membrane ‘shell’ of the peroxisomes is formed by building of the endoplasmic reticulum (ER), while the ‘content’ or matrix is imported from the cytosol (cytoplasmic matrix).
- However, there is now evidence suggesting that new peroxisomes always arise from pre-existing ones, being formed by growth and fission of old organelles similar to mitochondria and chloroplasts.
- Thus, peroxisomes are a collection of organelles with a constant membrane and a variable enzymatic content.
- All of their proteins (both structural and enzymatic) are encoded by nuclear genes and are synthesized in the cytosol (cytoplasmic matrix) (i.e., on the free ribosomes).
- The proteins present in either lumen or membrane of the peroxisome are taken up post-translationally from the cytosol (cytoplasmic matrix) as the haeme-free monomer;
- the monomers are imported into the lumen of peroxisomes,
- where they assemble into tetramers in the presence of haeme / Catalase and many peroxisomal proteins are found to have a signal sequence (comprising of three amino acids)
- which is located near their carboxyl ends and directs them to peroxisome .
- Peroxisomes contain receptors exposed on their cytosolic surface to recognize the signal on the imported proteins.
- All of the membrane proteins of the peroxisomes.
- Including signal receptor proteins, they are imported directly from the cytosol (cytoplasmic matrix).
- The lipids required to make new peroxisomal membranes are also imported from the cytosol (cytoplasmic matrix), possibly being carried by phospholipids transfer proteins from sites of their synthesis in the DR membranes.

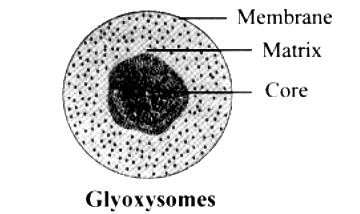
Glyoxysomes
- Glyoxysomes are found to occur in the cells of yeast,
- Neurospora, and oil rich seeds of many higher plants.
- They resemble with peroxisomes in morphological details, except that, their crystalloid core consists of dense rods of 6.0 mm diameter.
- They have enzymes for fatty acid metabolism and gluconeogenesis, i.e. conversion of stored lipid molecules of spherosomes of germinating seeds into the molecules of carbohydrates.
Functions
Glyoxysomes perform following biochemical activities of plants cells :
(1) Fatty acid metabolism.
- During germination of oily seeds, the stored lipid molecules of spherosomes are hydrolysed by the enzyme lipase (glycerol ester hydrolase) to glycerol and fatty acids.
- The phospholipids molecules are hydrolysed by the enzyme phospholipase.
- The long chain fatty acids which are released by the hydrolysis are then broken down by the successive removal of two carbon or C2 fragments in the process of b-oxidation.
- During b-oxidation process, the fatty acid is first activated by enzyme fatty acid thiokinase to fatty acyl-CoA which is oxidized by a FAD-linked enzyme fatty acyl-CoA dehydrogenase into-2-enoyl-CoA.
- Trans-2-enoyl-CoA is hydrated by an enzyme enoyl hydratase or crotonase to produce the L-3-hydroxyacyl-CoA, which is oxidized a NAD Linked L-3-hydratase or crotonase to produce the L-3-hydroxyacyl-CoA, which is oxidized by a NAD linked L-3-hydroxyacyl-CoA dehydrogenase to produce 3-Ketoacly-CoA.
- The 3-keto acyl-CoA looses a two carbon fragment under the action of the enzyme thiolase to generate an acetyl-CoA and a new fatty acyl-CoA with two less carbon atoms thatn the original.
- This new fatty acyl-CoA is then recycled thought the same series of reactions until the final two molecules of acetyl-CoA are produced.
- In plant seeds b-oxidation occurs in glyoxysomes .
- But in other plant cells b-oxidation occurs in glyxysomes and mitochondria.
- The glyoxysomal b-oxidation requires oxygen for oxidation of reduced flavorprotien produced as a result of the fatty-acyl-CoA dehydrogenase activity.
- In animal cells b-oxidation occurs in mitochondria.
- In plant cells, the acetyl-CoA, the product of b-oxidation chain is not oxidized by Krebs cycle, because it remains spatially separated from the enzymes of Krebs cycle, instead of it, acetyl-CoA undergoes the glyoxylate cycle to be converted into succinate.
(2) Glyoxylate cycle.
- The glyoxylate pathway occurs in glyoxysomes and it involve some of the reactions of the Krebs cycle in which citrate is formed from oxaloacetate and acetyl-CoA under the action of citrate synthetase enzymes.
- The citrate is subsequently converted into isocitrate by aconitase enzyme.
- The cycle then involves the enzymatic conversion of isocitrate to glyoxylate and succinate by isocitratase enzyme :
Isocitratase
Isocitrate Glyoxylate + Succinate
- The glyoxylate and another mole of acetyl-CoA form a mole of malate by malate synthesis ;
Malate synthetase
Acetyl CoA+Glyoxylate Malate
- This malate is converted to oxaloacetate by malate dehydrogenase for the cycle to be completed.
- Thus, overall, the glyoxylate pathway involves :
2 Acetyl-CoA+NAD+ Succintiate + NADH+ H+
- Succinate is the end product of the glyoxysomal metabolism of fatty acid and is not further metabolized within this organelle.
- The synthesis of hexose or gluconeogenesis involves the conversion of succinate to oxaloacetate, which presumably takes place in the mitochondria, since the glyoxsomes do not contain the enzymes fumarase and succinic dehydrogenase.
- Two molecules of oxaloacetate are formed from four molecules of acetyl-CoA without carbon loss.
- This oxaloacetate is converted to phosphoenol pyruvate in the phosphoenol pyruvate caboxykinase reaction with the loss of two molecules of CO2 :
2 Oxaloacete + 2ATP 2 Phosphoenol pyruvate + 2CO2 + 2ADP


